
神经干细胞的研究及其应用新进展
2025-10-14
999+
26.5KB
4 页
海报
侵权投诉
神经干细胞的研究及其应用新进展
[关键词] 神经干细胞 研究
健康讯:
崔桂萍 天津市脑系科中心医院 300060 1992 年, Reynolds 首次
成功地从成年小鼠纹状体中分离出神经干细胞( neural stem cell, NSC ),
于是“神经干细胞”这一概念被正式引入神经科学研究领域。可以总结为具有分
化为神经元、星形细胞和少突胶质细胞的能力,能自我更新并足以提供大量脑组
织细胞的细胞。不少文献中还提到神经祖细胞和神经前体细胞,目前认为,神经
祖细胞是指比 NSC 更有明确发展方向的细胞,而神经前体细胞是指处于发育早
期的增殖细胞,可指代 NSC 和神经祖细胞:与 NSC 相比,二者的分裂增殖能
力较弱而分化能力较强,是有限增殖细胞,但三者均属 NSC 范畴。 1. NSC 的
起源、存在部位及生物学特征 中枢神经系统的发育起源于神经沟、神经嵴、
神经管;研究发现, NSC 在神经管壁增殖,新生细胞呈放射状纤维迁移至脑的
特定位置;主要存在于室管膜区,在成脑生发区以外的区域也广泛分布,即具
有高度可塑性的神经前体细胞。 现发现 NSC 的生物学特征为:( 1 )具
有自我更新能力;( 2 )具有多向分化潜能,可分化为神经元、星形细胞和少
突胶质细胞;( 3 )处于高度未分化状态;( 4 )终生具有增殖分化能力,
在有损伤的局部环境信号变化的刺激下可以增殖分化。其中( 1 )和( 2 )是
NSC 的两个基本特征。 2. NSC 的基础研究进展 NSC 的增殖和分化调控是
目前 NSC 研究的核心问题,最近的研究资料显示, NSC 的增殖、分化、迁移调
控受多种相关因素的影响。 神经递质 神经递质作为细胞外环境的一员,
不仅介导神经元之间和神经元与效应器之间的信号传递,还参与 NSC 的增殖和
分化。这些神经递质包括谷氨酸( G1u )、 5- 羟色胺( 5-HT )、 GABA 、甘
氨酸( G1y )、乙酰胆碱( Ach )一氧化氮( NO )、肾上腺素与性激素等。
G1u :在脑的发育过程中有高含量的 G1u 表达, Haydar 等 发现, G1u 可
以通过大鼠胚胎皮质 AMPA/KAR 的激活调节室周区前体细胞的增殖,但 GLU 对
室管膜区( SZ )和室管膜下区( SVZ )体内细胞的影响是不同的,它可增加
SZ 细胞的增殖,减少 SVZ 细胞的增殖; GLU 还可促进神经元生长和分化。
5-HT :许多研究表明 , 5-HT 在皮质发育、突触形成中起重要作用,抑制 5-
HT 合成或选择性损伤 5-HT 神经元则引起齿状回及脑室下区神经元增殖活性下
降, 5-HT 可促进胶质细胞分化和髓鞘形成。 GABA : GABA 是成体脑发
育过程中主要的抑制性神经递质。 Haydar 等 发现, GABA 受体的激活可控制
神经前体细胞的细胞周期; Stewart 等 研究发现, GABA 和 G1u 对脑内不
同区域细胞增殖的影响是不同的,内源性 GABA 激活 GABA 受体在新皮质和调
节神经前体细胞增殖方面起重要作用。 G1y 及其它: G1y 受体( G1yR
)通过增加突触后细胞膜 C1 - 通透性而起突触后抑制作用。 Flint 等 发现,
G1yR 在胚胎大鼠和初生早期脊髓中为未成熟迁移和分化的神经元中起重要作
用,推测 G1yR 信号可能在突触形成中其重要作用; Ach 可通过 α -7 样烟
碱乙酰胆碱受体激活导致新生大鼠嗅球原代培养细胞神经突起过度生长,相反,
Ach 可抑制胚胎小鼠脊髓神经元的神经突起生长。有资料显示, NO 作为 CNS
的神经递质广泛参与神经细胞的存活、分化和可塑性的发生。而肾上腺素和性激
素则可使新生小鼠齿状回新生细胞数量减少。 细胞外基质 细胞外基质(
ECM )是组成间质和上皮血管中基质的不溶性结构成分,主要有胶原蛋白、弹性
蛋白、蛋白多糖和糖蛋白等。研究表明, ECM 可影响细胞分化、增殖、黏附、形态
发生和表型表达等生物学过程。 NSC 具有位置特异性的分化潜能,其增殖、分
化和迁移与 ECM 有非常密切的关系。 B- 链蛋白:新近资料表明, NSC
与 ECM 的黏附功能可以调节细胞的生长和增殖。 NSC 中的 B- 链蛋白和
Tcy/Lef 转录因子家族参与了细胞的成活、增殖和分化。 Chenn 等 发现,在
NSC 中稳定表达 B- 链蛋白的转基因小鼠,其发育的大脑皮质表面积增大,沟
回变深而宽,类似高级哺乳动物的皮质;侧脑室腔变大,与之相邻的脑室壁有
大量增生的细胞;并且其大部分 NSC 在有丝分裂后可重新进入细胞周期,说明
过度表达 B- 链蛋白并不破坏神经细胞正常发育分化,皮质的扩大是由于 NSC
增殖所致,提示 B- 链蛋白与 NSC 增殖有关。 Ree1in : Ree1in 是 ECM
中分子质量为 400 × 10 3 的蛋白质,与神经细胞表面的整合素受体 α 3 亚
基、极低密度脂蛋白和载脂蛋白 E 相结合,触发 Dab-1 胞液蛋白的衔接功能。
在皮质发育过程中的神经元以及脊髓节前神经元迁移中起重要作用。 细胞
黏附因子:细胞黏附因子是一种影响干细胞行为的重要信号蛋白,包括整合素
和黏合素等。研究表明, ECM 中的整合素在调控 NSC 增殖、分化和迁移方面有
重要的作用。脑内整合素与配体的相互作用促进了神经细胞的迁移,神经突起过
度生长和少突胶质细胞髓磷脂膜的形成,在可塑性过程的成体突触结构形成中
也起重要作用。黏合素家族中的 TN-C 在早期发育的中枢神经系统中广泛表达,
但在分化过程表达下降;成脑受伤后, TN-C 表达上调,提示 TN-C 在提高中
枢神经系统功能和可塑性方面有重要作用。 Garcion 等 用基因敲除 TN-C 的
方法,发现小鼠少突胶质前体细胞向视神经方向迁移增加,但在各脑区的增殖
率下降。 细胞生长因子: NSC 的增殖和分化还受多种细胞生长因子的调
控,如成纤维的细胞生长因子( FGF )和表皮生长因子( EGF )等。 FGF 有
三种受体, FGFR1 、 FGFR2 和 FGFR3 ,发育早期 FGF 在胎脑内进行增殖或神
经发生的区域内表达,成年脑内在相应的神经发生区内也有 FGF 的持续表达,
提示 FGF 在调节 NSC 增殖中发挥重要作用, EGF 在发育脑和成年脑内均有表
达,神经元和星形胶质细胞均可表达 EGF 。 糖蛋白:糖蛋白家族包括层
黏蛋白( LM ),纤维连接蛋白( FN )和腱蛋白( TN ), LM 为基底膜的
构成成分,可促进细胞黏附,调节细胞形态、分化及细胞迁移等; FN 具有形成
ECM ,促进细胞黏附、伸展、迁移、吞噬及血液凝固等多种生物学作用; TN 有
促进细胞黏附,促进或抑制细胞增殖和迁移等多种作用,并有拮抗 FN 的细胞
黏附作用。 Takano 等 新近发现, FN 对小鼠神经脊细胞中黑色素细胞的增殖
分化和迁移有重要作用。而 Chipperfield 等 则发现, ECM 中硫酸乙酰肝素
葡糖胺聚糖( HS )可促进 FGF-1 对成体 NSC 的有丝分裂作用。 基因调控
Notch 基因: Notch 信号通路对于决定胚胎发生、造血和 NSC 分化起着至关重
要的作用,当 Notch 被激活,干细胞进行增殖,当 Notch 活性被抑制,干细
胞进入分化程序,发育为功能细胞。 Tanigaki 等发现, Notch 在成体 NSC
发育为胶质细胞中起着重要作用,表达 Notch IC 明显增加星形细胞分化,减
少神经元和少突胶质细胞的产生。 bHLH 基因: bHLH 基因具有高度同源
性,是发育过程中转录络的重要组成部分,广泛参与神经和肌肉、细胞增殖分化
细胞谱系决定和性别决定等生理过程。 bHLH 基因在神经上皮细胞发育为神经元
中起关键并激活下游作用,可促进细胞脱离细胞周期,使细胞游离出皮质,并

摘要:
展开>>
收起<<
神经干细胞的研究及其应用新进展摘要1992年Reynolds首次从成年小鼠纹状体中分离出神经干细胞(NSC)“神经干细胞”概念正式进入神经科学研究领域。NSC具有分化为神经元、星形细胞和少突胶质细胞的能力并能自我更新。NSC的起源、存在部位及生物学特征中枢神经系统发育起源于神经沟、神经嵴、神经管NSC在神经管壁增殖并广泛分布于成脑生发区以外的区域。其生物学特征包括自我更新能力、多向分化潜能、高度未分化状态以及终生增殖分化能力。NSC的基础研究进展NSC的增殖和分化调控受神经递质、细胞外基质、细胞生长因子和基因调控等多种因素影响。其中神经递质如谷氨酸、5羟色胺、GABA等参与NSC的增殖和分化;细胞外基质通过B链蛋白、Reelin和细胞黏附因子等调节NSC行为;细胞生长因子如FGF和EGF对NSC增殖和分化有重要作用;基因调控方面Notch基因、bHLH基因、同源盒基因和Nestin基因等均参与NSC的发育和分化。NSC的应用研究进展随着对NSC了解的深入国内外科学家积极开展其临床应用研究包括细胞移植、基因载体治疗、神经损伤再生及生命科学研究。NSC移植可用于损伤神经细胞的替代基因载体治
相关推荐
-
妇科诊疗技术操作标准规范_20622084

 2025-11-29 999+
2025-11-29 999+ -
妇科诊疗常规(已审)_20625126

 2025-11-29 999+
2025-11-29 999+ -
妇科人流综合症应急预案及流程_20622076
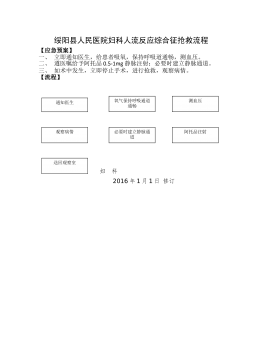 2025-11-29 999+
2025-11-29 999+ -
妇科检查及二合诊三合诊图解_20622068
 2025-11-29 999+
2025-11-29 999+ -
妇科检查规范_20622067
 2025-11-29 999+
2025-11-29 999+ -
妇科护理常规讲解_20625121
 2025-11-29 999+
2025-11-29 999+ -
妇科护理常规_20622061
 2025-11-29 999+
2025-11-29 999+ -
妇科 不孕病(多囊卵巢综合征)中医临床路径_20625117
 2025-11-29 999+
2025-11-29 999+ -
二级病原微生物试验室生物安全管理规范-上海卫生和计划生育_20621748
 2025-11-29 999+
2025-11-29 999+ -
2022子宫内膜异位症【29页】

 2025-11-29 999+
2025-11-29 999+

