
干细胞治疗糖尿病的研究近况
2025-10-14
999+
31.5KB
6 页
海报
侵权投诉
干细胞治疗糖尿病的研究近况
[关键词] 糖尿病 干细胞 胰岛
健康讯:
魏雪峰 史小林 100054 首都医科大学生殖医学研究中心 【摘
要】 本文对本世纪以来干细胞治疗糖尿病的若干研究,包括胰岛的体内发育、
分化及其标志物,以及应用胰导管干细胞和胚胎干细胞进行定向诱导分化为胰
岛对糖尿病进行细胞治疗的实验进行了综述。随着干细胞技术的不断完善,人类
最终能治愈糖尿病。 全球糖尿病(diabetes mellitus,DM)患者约亿
(我国也已达 4000 万),预计 2025 年将达 3 亿 [1],已成为严重危害人类
健康的重大疾病之一,特别是久病引发的多系统损害,及最终的肾功能衰竭。
DM 发病机制复杂,涉及遗传、环境、免疫调控和化学因子。自 1922 年发现了胰岛
素至今,DM 的治疗以药物和注射胰岛素为主,目前,学者们开始关注经胰岛移
植恢复患者体内代谢紊乱的方法,但因组织来源匮缺,制约了胰岛移植研究的
开展,因此寻找合适的胰岛移植物的研究便成为了当务之急。随着分子、细胞和
发育生物学技术的进展,增加了干细胞定向诱导分化成胰岛的可能性但由于这
一技术还处于实验室的实验阶段,而且方法也并不统一,因此有必要将近期研
究进行汇总,希望能对在这方面进行研究的同僚们有所帮助。 1 胰岛体内的
形成、发育、分化与鉴定 胚胎胰岛分化 胚胎胰岛的分化分 4 个阶段:(1)
胚胎 9~10 周为零散的多激素细胞阶段;(2)胚胎 11~15 周为未成熟的多激素
阶段;(3)胚胎 16~19 周为单激素核心胰岛阶段;(4)胚胎 30 周后为多形胰
岛阶段(Bocian,Histochem Cell Biol,1999)。在胰岛形成过程中,内胚层
细胞形成许多小导管,其末端膨大部分的细胞团发育为外分泌腺泡,与此同时,
一些细胞群或细胞索不出现管腔,卷曲成团并与其他细胞索分离开,分散在腺
泡之间,内含丰富的毛细血管,最后发育成具有内分泌功能的胰岛。 胰岛
细胞分化的调节 Ferber [2] 认为胚胎发育早期的信号源于脊索,后期的信
号源于间质,经诱导的胰管上皮细胞增殖分化形成胰岛细胞,但至今为止其诱
导因素并无统一认识。 PDX1:一种同源框转录因子(homeobox
tranˉscription factor),即胰十二指肠同源异型盒基因(pa-ncreatic and
duodenal homeobox1),编码胰腺十二指肠同源框蛋白 1,又称 IPF-1、IDX-
1、IUF-1。Lu(Lu,Endocrinology,1996)认为PDX1 是胰腺发育及胰岛素基因
转录表达的关键性转录因子,即决定于胰腺前体细胞向 B、A、D 细胞的分化 。
Mckinnon [3] 认为PDX1 对于肠内胚层背胰芽和腹胰芽的生长、分化起重要
作用。Dutta [4] 认为早期胰腺表达的 PDX-1 对胰腺上皮的形成和分化是必
需的。Gu [5] 认为所有胰腺细胞均来源于 PDX1表达的前体细胞,是最先发
现的胰腺发育决定基因,随着胰腺发育 PDX1 的表达逐渐减弱,如缺失将导致胰
腺无法形成。Docherty [6] 的研究证实小鼠 PDX1基因的纯合子缺失、突变会
导致胰腺无法形成。 ngn3:ngn3即神经元素 3(neurogenin3),是胰腺发育
过程中短暂表达的蛋白,可能是胰腺内分泌细胞系的定向因子,且在成熟胰岛
中不存在。用 PDX1启动子促进转基因小鼠中早期胰腺前体细胞中 ngn3表达,会
导致内分泌细胞数增加,外分泌细胞数减少。ngn3 缺失的纯合子小鼠中胰岛细
胞和 nestin+胰岛前体细胞在发育各阶段都缺失 [6] 。Lee [7] 认为
ngn3基因与胰岛分化有密切关系,因为 ngn3基因缺陷小鼠不能表达胰岛其他特
异性转录因子,包括 islet1、PAX4、PAX6,而缺乏 PAX6、NeuˉroD1、或的动物胰
腺中表达ngn3, 表明 ngn3位于这些因子的上游,且对启动内分泌细胞分化必
不可少,而分化后期关闭。Gasa [8] 通过表达的腺病毒感染胰腺导管细胞,
建立了体外调控分化模型,发现 ngn3和NeuroD1 活化胰岛分化的基因并不重叠,
表明这两个 bHLH 蛋白在胰岛发育活动中具有不同的功能。Mellitzer [9] 通
过将cDNA 插入到前体细胞的 3′端非翻译区,通过 EYFP 定位 NGN-3表达,发现
内分泌前体细胞在缺乏间充质信号刺激情况下,可在体外扩增,向内分泌细胞
分化为其默认的途径。Leon [10] 用抗新霉素基因筛选用含前体基因质粒转
染鼠胚胎干细胞。培养后用筛选阳性细胞,发现这些细胞分泌胰岛素,注入糖尿
病鼠体内可使其血糖恢复正常。 Foxα2:内分泌的关键调控基因是 forkhead
基因Foxα2。Guo [11] 诱导内胚层分化的研究发现,Foxα2 可被 Foxd3 激
活,而被 Oct4 抑制。内胚层分化可通过抑制Oct4 表达实现。Foxα2基因关键参
与肺、肝、胰发育。缺乏 Foxα2基因,鼠不能产生前、中肠内胚层
(Ang,Cell,1994)。 HNF:HNF4参与肝、肠、胰分化,缺失可致年轻人成熟
期发病的糖尿病(Maturity onset diabetes melliˉtus in
young,MODY1)。HNF1α(TCF1)HNF4或 Foxα2及Pdx-1双杂合缺失,导致 β
细胞功能缺陷,提示这些基因构成β细胞发育的基因络
(Yamagata,Nature,1996)。HNF6-/-鼠,腹胰发育缺陷及胆管发育异常,该
基因需 ngn3 激活(Yamagata,Nature,1996)。HNF6 还激活 Foxα2a 基因。
Pax:Pax-4和Pax-6 同属 Pax 基因家族,其在胰腺内分泌细胞分化中起关键作用。
Sosa(Sosa,Nature,1997)将 Lac-Z 插入到 Pax 基因中作标记,发现 Pax-4
失活的新生鼠经免疫组织化学法显示缺少β细胞和 δ细胞,α细胞排列不规
整。Pax-6 缺失,α细胞缺如,而胰岛素和生长抑素分泌细胞却可出现(St-
Onge,Nature,997)。由此推断出 Pax-6 是α细胞发育所必需,而非 β、δ细
胞发育所必需。 Chiang [12] 为分析单个胰腺细胞的转录模式,改进了
基于PCR 的cDNA 扩增方法,即将单个胰腺细胞中的 cDNA 与含有 95 个常见胰腺
细胞表达基因的 DNA 芯片进行杂交,芯片上的基因包括转录因子,信号传导分
子以及对照基因。研究者观察了胚胎小鼠的胰腺分离出的单个细胞中的基因表达
模式。在这个时期上皮细胞的形态均一,而基因表达模式分析可分为 6个不同的
亚型。I 型只表达Pdx1,和,II 型还表达P48,III 型还表达ngn3。因为 Pdx1 是
最早表达的胰腺脏细胞标记蛋白,因此研究者认为I 型细胞是最好的胰腺干细
胞候选者。II 型细胞表达的 P48是外分泌胰腺细胞的标记,而和是内分泌细胞
分化所必须的。研究者根据这些研究结果推测出一个胰腺细胞分化的模型,每一
个阶段都对应相应的基因表达模式。此外,成体的胰岛内分泌细胞 40~50d 更新
一次,其过程包括细胞凋亡和胰导管干细胞增殖分化成新的胰岛细胞。给予葡萄
糖或高血糖素样肽(Glucagon-like peptied,GLP1)等促胰岛因子 48h 后,胰
岛细胞数量可增加一倍,说明机体胰岛在一定的条件与需求下进行着不断更新
与增殖的(Rosenberg,Cell Transplant,1995)。 胰岛前体细胞的标记
Jindal(Jindal,Transplanˉtation Proceeding,1997)认为胰脏的外分泌
细胞和内分泌细胞都来源于具有导管细胞特征的未分化上皮细胞,提示胰导管
细胞可能是胰岛的前体细胞。Berger [13] 也发现胰腺中存在干细胞,认为
胰腺干细胞是胚胎发生中没有进行终末分化的胰腺细胞,具有自我更新和分裂
能力。应用 HE 染色显示该细胞是一种嗜碱性的单核细胞,直径约8μm,呈圆形,

标签: #糖尿病
摘要:
展开>>
收起<<
摘要本文综述了本世纪以来干细胞治疗糖尿病的研究进展包括胰岛的体内发育、分化及其标志物以及利用胰导管干细胞和胚胎干细胞定向诱导分化为胰岛对糖尿病进行细胞治疗的实验。全球糖尿病患者数量庞大DM发病机制复杂目前治疗以药物和注射胰岛素为主但胰岛移植因组织来源匮缺而受限干细胞定向诱导分化成胰岛成为研究热点。在胰岛形成过程中内胚层细胞形成小导管部分细胞团发育为外分泌腺泡一些细胞群发育成具有内分泌功能的胰岛。胰岛细胞分化受多种因素调节如PDX1是胰腺发育及胰岛素基因转录表达的关键性转录因子ngn3与胰岛分化密切相关。近年研究揭示了胰岛前体细胞的特异性标记物如酪氨酸羟化酶、葡萄糖转运子2等。诱导干细胞分化为胰岛的研究中胰导管干细胞和胚胎干细胞是主要来源但将这些细胞诱导成胰岛仅处于实验室研究初级阶段。
相关推荐
-
妇科诊疗技术操作标准规范_20622084

 2025-11-29 999+
2025-11-29 999+ -
妇科诊疗常规(已审)_20625126

 2025-11-29 999+
2025-11-29 999+ -
妇科人流综合症应急预案及流程_20622076
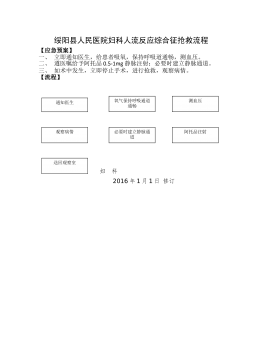 2025-11-29 999+
2025-11-29 999+ -
妇科检查及二合诊三合诊图解_20622068
 2025-11-29 999+
2025-11-29 999+ -
妇科检查规范_20622067
 2025-11-29 999+
2025-11-29 999+ -
妇科护理常规讲解_20625121
 2025-11-29 999+
2025-11-29 999+ -
妇科护理常规_20622061
 2025-11-29 999+
2025-11-29 999+ -
妇科 不孕病(多囊卵巢综合征)中医临床路径_20625117
 2025-11-29 999+
2025-11-29 999+ -
二级病原微生物试验室生物安全管理规范-上海卫生和计划生育_20621748
 2025-11-29 999+
2025-11-29 999+ -
2022子宫内膜异位症【29页】

 2025-11-29 999+
2025-11-29 999+

